Microbiome as a Source of B Vitamins
by Mary Ferrari
“Our gut harbors bacteria that produce B vitamins, including biotin, cobalamin, folate, niacin, pantothenate, pyridoxine, riboflavin, and thiamine.”
B vitamins are a group of water-soluble nutrients that play essential roles in nearly every aspect of human metabolism that function primarily as coenzymes, helping the body carry out hundreds of biochemical reactions involved in energy production, DNA synthesis, nervous system function, and immune regulation. Although these vitamins are traditionally associated with preventing deficiency diseases, recent research has revealed that they also have important effects on gut health and the composition of the intestinal microbiome. The relationship between B vitamins and the gut is complex because these nutrients not only influence the host but also affect the growth, metabolism, and interactions of the trillions of microorganisms that reside in the digestive tract.
The human gut microbiome consists of a vast community of bacteria, fungi, viruses, and other microorganisms that contribute to digestion, immune development, and overall health. Many members of this microbial community can synthesize B vitamins, while others depend on obtaining them from their environment creating an intricate network of nutrient exchange within the gut ecosystem. Some bacterial species produce vitamins that can be used by neighboring microbes, while other species compete for the same nutrients and as a result, the availability of B vitamins can influence which microbial populations thrive and which decline. These interactions help shape the overall composition and stability of the gut microbiota.
Several gut bacteria are capable of producing important B vitamins, including thiamine (B1), riboflavin (B2), niacin (B3), pantothenic acid (B5), pyridoxine (B6), biotin (B7), folate (B9), and cobalamin (B12) but not all bacteria possess the genetic machinery required to synthesize every vitamin. Some species are self-sufficient, whereas others rely on vitamins produced by neighboring microbes or supplied through the diet. This interdependence creates a highly connected microbial community in which nutrient availability can influence bacterial diversity and ecological balance. Researchers have also found that many beneficial gut bacteria participate in vitamin production highlighting another way in which the microbiome contributes to host health.
Among the B vitamins, folate and vitamin B12 are particularly important because they participate in one-carbon metabolism, a series of biochemical reactions required for DNA synthesis and cellular growth. Deficiencies in these vitamins can impair cell division and affect rapidly growing tissues, including the intestinal lining and because the cells lining the gut are constantly renewed, an adequate supply of these nutrients is essential for maintaining intestinal integrity. When deficiencies occur, the barrier function of the intestine may become compromised increasing susceptibility to inflammation and disease.
B vitamins also play significant roles in maintaining immune function. Several members of the vitamin B family contribute to the development and activity of immune cells that help protect the body from infection. For example, vitamins B6, B9, and B12 are involved in the production and maturation of lymphocytes, which are essential components of the adaptive immune system. Insufficient levels of these vitamins may impair immune responses and contribute to chronic inflammation. Since the gut contains a large proportion of the body’s immune cells, disruptions in vitamin availability can have important consequences for intestinal health and immune regulation.
Research has demonstrated that deficiencies in B vitamins can affect the structure and function of the gastrointestinal tract. Inadequate levels may alter intestinal morphology, reduce the efficiency of nutrient absorption, and impair the maintenance of the epithelial barrier that separates gut microbes from underlying tissues. Damage to this barrier can promote inflammation and increase the risk of gastrointestinal disorders. Studies have linked deficiencies in various B vitamins to conditions characterized by intestinal dysfunction, highlighting the importance of adequate vitamin intake for maintaining a healthy digestive system.
The relationship between B vitamins and the microbiome is bidirectional. While vitamins influence microbial growth, gut microbes can also affect the host’s vitamin status. Certain bacterial species contribute significantly to the production of vitamins that may be absorbed and utilized by the host. Changes in the composition of the microbiome, whether due to diet, antibiotic use, illness, or other factors, can therefore alter vitamin production within the intestine. This has led researchers to investigate whether manipulating the microbiome through probiotics, prebiotics, or dietary interventions could enhance vitamin synthesis and improve health outcomes.
Emerging evidence suggests that disruptions in vitamin-microbiome interactions may contribute to a variety of gastrointestinal disorders. Conditions such as inflammatory bowel disease, irritable bowel syndrome, and other inflammatory disorders are often associated with altered microbial communities and nutritional imbalances. Although the exact mechanisms remain under investigation, researchers surmise that deficiencies in certain B vitamins may exacerbate intestinal inflammation and impair the body’s ability to maintain a healthy gut environment. Conversely, restoring adequate vitamin levels may help support intestinal healing and promote microbial balance.
The growing understanding of B vitamins and gut health has important implications for nutrition and disease prevention. Rather than viewing vitamins solely as dietary requirements for the host, scientists now recognize them as key factors influencing the ecology of the gut microbiome. These nutrients help regulate microbial interactions, support intestinal barrier function, contribute to immune health, and participate in numerous metabolic pathways essential for both host and microbial survival. As research continues, a deeper understanding of vitamin-microbiome relationships may lead to new strategies for improving digestive health through personalized nutrition and microbiome-targeted therapies.
In summary, B vitamins serve far more functions than simply preventing deficiency diseases. They are integral components of a dynamic relationship between the host and the gut microbiota. Through their effects on metabolism, immune regulation, microbial ecology, and intestinal integrity, B vitamins play a central role in maintaining gut health and overall well-being. Continued research in this area is expanding our understanding of how nutrition and the microbiome work together to support human health throughout life.
Vitamin B1/Thiamine
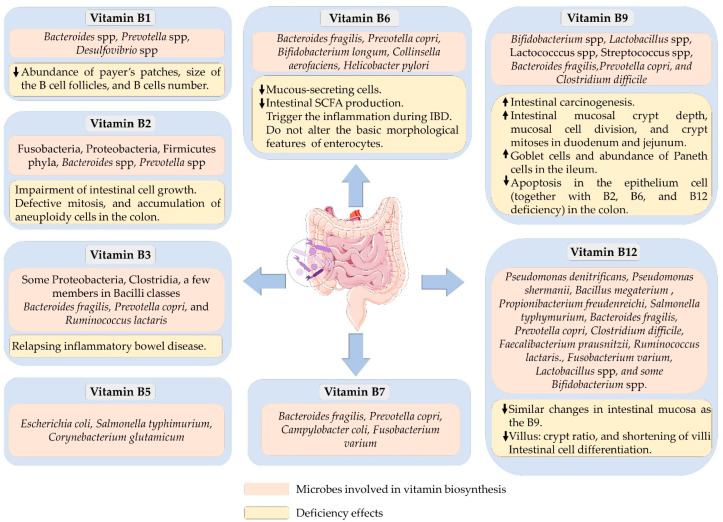
Thiamine is an essential cofactor required for several enzymes, especially in glycolysis, the tricarboxylic acid (TCA) cycle, and the pentose phosphate pathway. Several bacteria can synthesize free thiamine or its active form, thiamine pyrophosphate (TPP). However, bacteria belonging to the enterotype 2 class have higher thiamine biosynthetic capacity. This class of bacteria, mainly Prevotella and Desulfovibrio, is overexpressed with four enzymes—hydroxymethylpyridine kinase, phosphomethylpyridine kinase, thiamine-phosphate pyrophosphorylase, and thiamine-monophosphate kinase—crucial for thiamin biosynthesis. Additionally, more than 90% of other gut-associated Bacteroides possess the ability for thiamin biosynthesis and the thiamin transporter genes. Dietary free thiamine is absorbed through carrier-mediated transportation such as high-affinity thiamin transporter 1 (THTR-1) and 2 (THTR-2) located in the epithelium and mucosa of the small intestine. If the dietary thiamine is in the bound form, such as TPP, they have to be first converted to free thiamin before it is absorbed. Likewise, the above transporters, i.e., THTR-1 and THTR_2, are primarily responsible for the absorption of bacterially produced free thiamin in the colon. However, TPP produced by gut microbiota is absorbed directly by TPP transporters such as TPPT -1, indicating a difference in dietary and bacterially produced TPP absorption. Although the role of thiamine on intestinal integrity is not well understood, some have suggested having certain gut-related immune regulatory functions through a recent concept involving energy metabolism. Mathis and Shoelson suggested that thiamine directs the energy balance between glycolysis and TCA cycle activities, controlling immunometabolism. It is also likely to have some roles associated with intestinal-linked immune cells. It has been shown that thiamine deficiency reduces the abundance of the Payer’s patches and decreases the size of B-cell follicles, leading to a reduction in naïve B cells in female Balb/c mice. Their findings indicated a potential gut-related role of thiamine in immunometabolism.
Vitamin B2/Riboflavin
Riboflavin is another micronutrient involved in the energy-producing reactions of carbohydrate, fat, and protein metabolisms. Riboflavin is converted to its active forms, flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD), by flavokinase and FAD synthetase, respectively. Human gut microbiota can produce riboflavin, primarily in the large intestine. Dietary riboflavin ingested in FAD or FMN forms must be converted into free riboflavin before absorption. In humans, riboflavin absorption occurs mainly in the proximal small intestine through active carrier-mediated transportation. A complete riboflavin operon is present in all Baceteroidetes and Fusobacteria, as well as in 92% of Proteobacteria, indicating they are the primary riboflavin producers in the gut. In addition, half of the Firmicutes are predicted to be riboflavin producers. Lactic acid bacteria from dairy products also possess riboflavin biosynthesis capacity. These bacteria synthesize riboflavin by utilizing guanosin-5’-triphosphate (GTP), a compound derived from the purine biosynthesis pathway, and ribulose-5-phosphate, an intermediate from the pentose phosphate pathway.
Interestingly, riboflavin is also required for the postnatal development of the gastrointestinal tract. Its deficiency was associated with crypt hypertrophy, interruptions in crypt bifurcation in rats, and loss of proliferative potentials in intestinal cells. These changes were seen in the postnatal and post-weaning stages. The changes were irreversible even after the repletion of riboflavin in both in vivo and in vitro experiments. Williams and colleagues have shown that riboflavin deficiency reduced villus number but increased its length. Riboflavin depletion in humans was also associated with shorter duodenal crypt and low cell division. In vitro studies using Caco-2, HCT116, and HT29 cells demonstrated potential mechanisms of riboflavin deficiency phenotype. They include inhibiting cell growth by reducing cellular ATP generation and elevating oxidative stress, defective mitosis, and accumulation of aneuploidy cells. The changes in this intestinal morphology might also be related to its adaptive responses to deficiency-related stresses. On the other hand, riboflavin supplementation increased the abundance of bacteria that cannot synthesize riboflavin, such as Faecalibacterium prausnitzii and Roseburia spp.. Higher intake was also associated with an increase in the abundance of Prevotella spp. but Bacteroides’ concentration was decreased in lactating women.
Vitamin B3/Niacin
Vitamin B3, known as nicotinamide, nicotinic acid, or niacin, is converted into its active form, nicotinamide adenine dinucleotide (NAD), which is essential for many critical metabolic processes, primarily as a redox cofactor. Similar to other higher organisms, intestinal bacteria synthesize vitamin B3 mainly from amino acid tryptophan but using a unique pathway. According to the genetic assessment, the Bacilli class contains only 4, the Clostridia class contains 44, and Proteobacteria contains 29 potential niacin synthesizing bacteria. Bacteroides fragilis, Prevotella copri, and Ruminococcus lactaris can also produce vitamin B3 in the gut as they possess a vitamin B3 biosynthesis pathway. Moreover, niacin-responsive transcription factor NiaR (YrxA) is present in a diverse group of Bacillus and Clostridium bacteria, meaning they can undergo de novo synthesis of NAD. The human and mouse colonic epithelial cells possess an efficient, specific, and regulated mechanism for the uptake of vitamin B3. The bacterially synthesized vitamin B3 contributes to local colonocyte nutrition and maintains the morphology of intestinal stem cells.
A study from the FoCus cohort identified links between vitamin B3 and intestinal microbial composition. They found a significant association between vitamin B3 deficiency and low α-diversity and the abundance of Bacteroidetes in obese individuals. The abundance of this bacteria was significantly higher during a gut-targeted delayed-release of nicotinic acid but not the nicotinamide in those obese subjects. Qi and colleagues isolated intestinal crypt cells from a C3H/HeN conventionally raised mouse and treated them with vitamin B3. Vitamin B3 treatment with 1200 ug/mL significantly increased the growth rate of organoids. Vitamin B3 plays a vital role in reducing inflammation and causes relapsing inflammatory bowel diseases such as ulcerative colitis when deficient. It controls inflammation by inhibiting vascular permeability in intestinal tissues by activating the PGD2/DP1 signal in endothelial cells. It also modulated the inflammatory response by enhancing the rate of ATP generation in Caco-2 cells. Interestingly, vitamin B3 engages in various metabolic reactions that alter the cellular redox state and rapamycin signalling pathway, thus suppressing the colon’s inflammations.
Furthermore, vitamin B3 protects colonic epithelial cells against the dextran-sulfate-sodium (DSS)-induced apoptosis and promotes cell proliferation in mice. It maintains the intestinal epithelium barrier by activating the D prostanoid 1 (DP1) receptor in macrophages and endothelial and colonic epithelial cells. Lower plasma niacin levels have been observed in patients with Crohn’s disease. Interestingly, retention enema containing vitamin B3 effectively promoted mucosal healing in patients with ulcerative colitis, most likely due to the downregulation of colonic inflammatory cytokines and suppression of proinflammatory gene expression. In another in vitro study, cellular metabolites such as glutamine, isoleucine, ornithine, and glycerophosphocholine were downregulated, and glutamic acid was upregulated in inflamed Caco-2 cells. The impairments in the metabolite profile were ameliorated with the addition of vitamin B3, which means that the therapeutic properties of vitamin B3 might be related to improving specific cellular metabolites that were impaired during acute inflammations.
Vitamin B5/Pantothenic Acid
Vitamin B5, or pantothenic acid, is an essential precursor of coenzyme A (CoA) and acts as an acyl-carrier protein. It is involved in various metabolic pathways such as the citric acid cycle, cell growth, neurotransmitter synthesis, and fatty acid oxidation. Various bacteria, including Escherichia coli, Salmonella typhimurium, and Corynebacterium glutamicum, can synthesize vitamin B5. They use aspartate and intermediate metabolites of valine biosynthesis to produce vitamin B5. For instance, S. Typhimurium produces pantothenate from α-ketoisovalerate using acetohydroxy acid synthase isozyme I and dihydroxy acid dehydratase enzymes. Moreover, other bacteria, such as Lactobacillus helveticus, require pantothenic acid for their fatty acid and biotin metabolism. Dietary pantothenic acid supplementation also influences gut microbial profile. Enhanced pantothenic acid intake increased the relative abundance of Prevotella and Actinobacteria and decreased the abundance of Bacteroides in lactating women.
Vitamin B6/Pyridoxine
Vitamin B6 has six vitamers, namely, pyridoxine (PN); pyridoxal (PL); pyridoxamine (PM); and their phosphorylated forms, i.e., pyridoxal phosphate (PLP), pyridoxine phosphate (PNP), and pyridoxamine phosphate (PMP) [64]. PLP is the enzymatically most active form of vitamin B6. Vitamin B6 acts as a cofactor for many biochemical reactions, primarily involved in amino acid biosynthesis and catabolism. Besides this, it is involved in fatty acid and neurotransmitter biosynthesis and also acts as an antioxidant. In the mammalian gut, bacteria synthesize vitamin B6 through de novo or salvage pathways. Microbes such as Bacteroides fragilis and Prevotella copri (Bacteroidetes), Bifidobacterium longum and Collinsella aerofaciens (Actinobacteria), and Helicobacter pylori (Proteobacteria) can produce vitamin B6 as they have these biosynthetic mechanisms.
Most dietary vitamins are absorbed in the small intestine; however, uptake of a certain amount of dietary and bacterially synthesized vitamin B6 still occurs in the large intestine because many vitamin B6 transporters are also expressed in the mammalian colon. Vitamin B6 auxotrophic prokaryotes and single-cell eukaryotes rely on importing this vitamin from their surroundings, while multicellular organisms transport it to different host organs after absorption. Vitamin B6 produced in the gut is not sufficient for the host’s daily requirements. Its deficiency reduced microbial β-diversity and significantly altered intestinal metabolite compared to the control groups in rats [70]. An abundance of Lachnospiraceae_NK4A136_group and Prevotella were elevated with vitamin B6 deficiency, while the abundance of Bacteroides was decreased when the vitamin B6 intake was high. Moreover, Bifidobacterium, Slackia, Enterococcus, Thiococcus, Klebsiella, Serratia, and Enterobacter abundances were also decreased with vitamin B6 supplementation in lactose-intolerant patients.
A cross-sectional study has shown an association of severity of irritable bowel disease symptoms with low dietary vitamin B6 intake. A plausible explanation includes the trigger of inflammation by shifting the balance between anti-inflammatory to proinflammatory cytokines with low vitamin B6. The presence of a P2X receptor antagonist such as pyridoxal phosphate 6-azophenyl-2,4-disulfonic acid, a derivative of vitamin B6, and impairment in microbiota-related intestinal metabolites such as short-chain fatty acids might play a significant role in triggering inflammation. Recently, Yin and colleagues demonstrated that dietary supplementation of vitamin B6 downregulated the inflammatory cytokines and upregulated the mRNA expression of amino acid transporters in the jejunum of weaned piglets. Moreover, vitamin B6 deprivation studies on aquatic animals have shown a significant decrease in the number of mucous-secreting cells, a critical factor in maintaining gut health. Nevertheless, vitamin B6 deficiency did not alter the basic morphological features of enterocytes, such as cell viability, cell volume, membrane permeability, and protein content in rats, but decreased calcium transport flux.
Vitamin B7/Biotin
Vitamin B7 acts as a coenzyme for several biochemical reactions, such as glycolysis and cell signalling and epigenetic regulations. It also controls gene expression, including nuclear factor kappa B (NF-κB), through histone binding mechanism, commonly known as biotinylation. Therefore, this vitamin may also have anti-inflammatory effects. Biotin is primarily synthesized from either malonyl CoA or pimeloyl-CoA. Enzymes of the biotin biosynthesis pathway are overrepresented in enterotype 1, enriched in Bacteroides. Bacteria that can produce vitamin B7 include Bacteroides fragilis, Prevotella copri, Fusobacterium varium, and Campylobacter coli. In contrast, others are extensive vitamin B7 reducers, such as Lactobacillus murinus.
Vitamin B9/Folate
Folate, also known as vitamin B9, is a conjugate form of 4-aminobenzoic acid and L-glutamic acid. Vitamin B9 is supplied to the host primarily through diet and partially by gut microbiota. Folate is an essential methyl donor nutrient that provides one-carbon units. It is also involved in synthesizing S-adenosylmethionine (SAM) required for cellular biosynthesis and DNA methylation. This vitamin is vital for replicating and restoring nucleic acids, thus affecting cell survival rate and proliferation when deficient. Additionally, folate is involved in regulating gene activities, regenerating the lining of the intestine, producing necessary chemicals for proper brain function, decreasing the growth of lymphocytes, and reducing natural killer cell cytotoxicity. Thus, every living cell requires folate to perform a variety of these biochemical and biosynthetic processes. These cellular reactions are universal, but their metabolic pathways differ from organism to organism. Organisms such as fungi, plants, bacteria, and some specific archaea can undergo folate biosynthesis, and they use a similar pathway with slight modifications. There are several bacteria that can produce folate in the gut, which include Bacteroides fragilis, Prevotella copri, Clostridium difficile, Lactobacillus Plantarum, L. reuteri, L. delbrueckii ssp. bulgaricus, Streptococcus thermophilus, Bifidobacterium spp. (some species), Fusobacterium varium, and Salmonella enterica. Out of these bacteria, Bifidobacterium species are well studied. They are categorized on the basis of their folate-producing ability: high folate producers—Bifidobacterium bifidum and B. longum subsp. Infantis, and low folate producers—B. breve, B. longum subsp. longum, and B. adolescentis. Synthesis of folate requires one pterin moiety originating from 6-hydroxymethyl-7,8-dihydropterin pyrophosphate (DHPPP) and a para-aminobenzoic acid (pABA). The latter is an intermediate formed by cleaving pyruvate with 4-amino-4-deoxychorismate lyase enzyme [98]. Such enzymes are mostly confined to the genomes of Bifidobacterium species, including B. adolescentis and B. dentium Bd1; thus, they produce folate when DHPPP is available. Other common folate-producing bacteria are Lactobacilli. Unlike Bifidobacterium, several Lactobacillus species, including L. plantarum, L. sakei, L. delbrueckii, L. reuteri, L. helveticus, and L. fermentum can produce DHPPP; therefore, they synthesize folate when pABA is available. Nevertheless, Lactococcus and Streptococcus possess a complete pathway for the de novo folate biosynthesis and do not require a supply of either DHPPP or pABA.
Due to its crucial role in methyl donor production, folate deficiency significantly impairs DNA replication. Folate depletion causes an increase in the intestinal mucosal crypt depth in the duodenum and jejunum, resulting in a reduced villus to crypt ratio. In methyl donor-deficient mice induced by feeding a folate-deficient diet accompanied by an antibiotic, succinylsulfathiazole (1%), also had increased crypt depth and altered intestinal cell differentiation. In rats, folate deficiency causes megaloblastic changes in the epithelial cell nuclei and reduced crypt mitosis. These changes were more remarkable in the ileum with crypt elongation, increased goblet cells, and decreased Paneth cells. Thus, folate deficiency significantly alters intestinal cell morphology and is associated with increased occurrence of intestinal carcinogenesis. Concomitant depletion of folate, riboflavin, vitamin B-6, and vitamin B-12 alter Wnt- signalling in the mouse colon and decrease apoptosis in the epithelium cell. Unexpectedly, these changes are irreversible, even with the repletion of folate. Although the gut bacteria can produce some folate, a folate-deficient diet significantly alters microbial diversity in the mice. It is shown that the abundances of Bacteroidales and Clostridiales decreased, and abundances of Lactobacillales and Erysipelotrichaceae taxa increased in folate-deficient mice.
Vitamin B12/Cobalamin
Vitamin B12, also known as cobalamin, is one of the largest and most complex vitamins. The other forms of this vitamin include cyano-, methyl-, deoxyadenosyl-, and hydroxy-cobalamin. The cyano form is found as traces in diet, and it is used as a dietary supplement. Like folate, cobalamin is involved in methyl donor synthesis, such as SAM. These methyl donors are crucial for nucleic acid synthesis and protein and lipid metabolism. It is used as a cofactor for methionine synthase in sulphur amino acid metabolism to recycle homocysteine to methionine. Cobalamin is also vital for the proper functioning of the central nervous system and the synthesis of red blood cells.
There are limited bacteria that can synthesize vitamin B12 in the human gut, and most of them use precorrin-2 as a precursor. Approximately 20% of gut bacteria can produce vitamin B12, and more than 80% of gut bacteria require B12 for their metabolic reactions. These include Pseudomonas denitrificans, Bacillus megaterium, and Propionibacterium freudenreichi, Bacteroides fragilis, Prevotella copri, Clostridium difficile, Faecalibacterium prausnitzii, Ruminococcus lactaris, Bifidobacterium animalis, B.infantis, B.longum, and Fusobacterium varium. Notably, the first three bacteria are commercially used for vitamin B12 production. Biosynthesis of B12 by microorganisms involves almost 30 genes and uses either aerobic or anaerobic pathways. The aerobic pathway has been studied in Pseudomonas denitrificans, and the anaerobic pathway has been studied in Salmonella typhimurium, Bacillus megaterium, and P. shermanii. In general, Lactobacillus spp. were thought not to have a vitamin B12 biosynthetic pathway. However, the discovery of the conversion of glycerol into propanediol in lactic acid bacteria demonstrated their ability to produce vitamin B12.
On the other hand, several other bacteria, including Bacteroides, do not have a vitamin B12 biosynthesis capability. However, most of them possess vitamin-B12-dependent enzymes. The optimum functioning of these enzymes depends on dietary supply. The effects of vitamin B12 deficiency on colon morphology are similar to that of folate deficiency as they are closely associated with several cellular metabolic reactions. However, the impact of its deficiency on colon inflammation is not conclusive. Benight and colleagues reported that vitamin B12 deficiency protects against DSS-induced inflammations in C57BL/6 mice. Contrarily, others demonstrated a reduction in cell differentiation and intestinal barrier in vitamin-B12-deficient rats. Moreover, in patients with a vitamin B12 deficiency, the villus becomes shorter with a reduced villus/crypt ratio than in the control group. Similarly, a dietary deficiency or surplus of vitamin B12 may likely influence the growth of gut microbiota. Unexpectedly, vitamin B12 deficiency did not alter gut microbial composition in healthy mice but altered it in DSS-induced colitis mice. The short time (28 days) applied to induce the deficiency might be one of the reasons that gut microbial composition was not affected in the healthy mice. It was also likely that the animals that practice coprophagy could have maintained their vitamin status by eating feces. However, the gut microbial profile in humans is influenced by the host vitamin B12 status. Vitamin B12 supplementation in humans increased the relative abundance of Prevotella, but decreased the abundance of Bacteroides. Likewise, the relative abundance of Bacteroides has reduced with vitamin B12 supplementation in C57BL/6 mice. Lurz and colleagues also showed that vitamin B12 supplementation in mice significantly decreased Parabacteroides and Lactobacillus and increased E. coli and Enterococcus abundances in a murine model of colitis.

Immune For Life
Forgotten factors for life and health that are shaping the future of drugs and food.
Journey to a healthier lifestyle

Immune For Life
M. Ferrari
After decades of chronic health conditions and serious gut issues like IBS and SIBO, immune deficiencies and an autoimmune condition discover how I recovered my health thanks to natural oral immune therapeutics (maf and gcmaf). Due to a premature birth and being formula fed, I was a SAM child in real life. My book is a step by step journey you won't want to miss that illustrates how to regain or maintain health for all ages.